

Introdução
Os porcos, num sistema de produção comercial, estão continuamente expostos a vários agentes patogénicos e vírus. A exposição crónica a infecções clínicas ou subclínicas compromete a função da barreira intestinal e ativa uma resposta sistémica que reduz a taxa de deposição de proteínas e a eficiência do crescimento. A resposta sistémica afeta o rendimento dos porcos de duas maneiras: altera a partição de nutrientes e produz respostas neurológicas à infecção induzidas por eicosanoides (por exemplo, anorexia). Neste artigo, far-se-á uma revisão do papel da nutrição como atenuante da redução do rendimento causado pela ativação do sistema imunitário.

Cumprir com a alteração das necessidades de aminoácidos
A ativação do sistema imunológico reduz a eficiência alimentar ao alterar a partição de nutrientes. A libertação de citocinas pró e anti-inflamatórias estimula o fígado e aumenta a captação hepática de aminoácidos para produzir moléculas relacionadas com a imunidade (Breuille et al., 1994; 1998). Na musculatura esquelética de porcos e ratos com o sistema imunitário ativo, é evidente um significativo catabolismo de aminoácidos, que depende da gravidade e duração da inflamação (Breuille et al., 1994; 1998; Williams et al., 1997a; 1997b). Rakhshandeh e de Lange, 2011, publicaram uma revisão exaustiva sobre o papel da cada aminoácido na deposição total de proteína em porcos com o sistema imunológico ativado. Em sintese, apesar de um aumento na procura metabólica de glutamina, arginina, fenilalanina, tirosina e de aminoácidos de cadeia ramificada durante uma ativação moderada do sistema imunitário, as necessidades não aumentaram por causa da sintese de novo ou porque a disponibilidade na dieta era suficiente para cobri-los. Embora tenhamos uma informação muito limitada, acredita-se que as necessidades de treonina podem aumentar quando se estimula a produção de muco ou de imunoglobulinas ricas em treonina durante a ativação do sistema imunitário porque a treonina é o principal aminoácido do muco e de algumas imunoglobulinas (Faure et al., 2007; Rakhshandeh e de Lange, 2011). Utilizando uma injeção intramuscular de um lipopolissacarídeo (LPS) de E. coli, demonstrou-se que as necessidades de triptofano cresceram 7% em porcos em crescimento com o sistema imunitário ativo (de Ridder et al., 2012). É sabido que as necessidades em aminoácidos sulfurados (AAS), especialmente em cisteína, aumentam nos porcos com o sistema imunológico ativo. Recentemente tem havido estudos em leitões desmamados e porcos de engorda com modelos de infeção por E. coli e por injecção de LPS, respetivamente, para reproduzir o nível de ativação imunológica de explorações comerciais, onde se produzem desafios crónicos por agentes patogénicos e vírus (ver figuras 1 e 2). Estes estudos demostraram que, em comparação com os porcos saudáveis, a taxa de deposição total de proteínas nos que têm o sitema imunológico ativado decresceu 12% em dietas com os valores recomendados para AAS (NRC, 2012) e a deposição proteica máxima só se atingiu com a suplementação de AAS, 20% acima das recomendações atuais da NRC (relação SAA:Lys de 0,75; Kim et al., 2012).
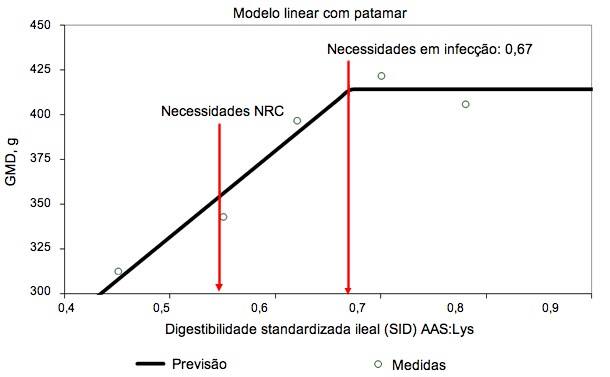
Figura 1. Necessidades de digestibilidade ileal standardizada para aminoácidos sulfurados, determinada em leitões desmamados infetados oralmente com uma estirpe enterotoxigénica de E. coli (Capozzalo et al., dados não publicados)
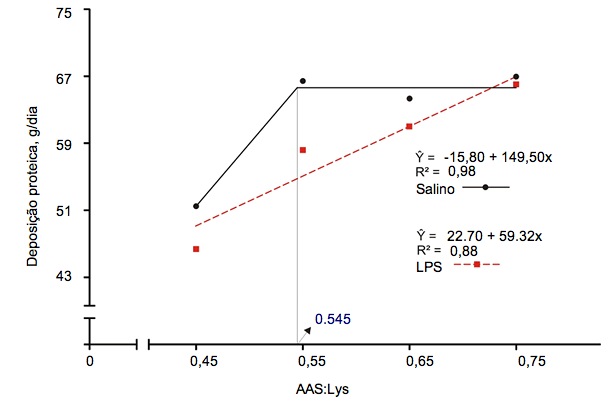
Figura 2. Necessidades de digestibilidade ileal standardizada para aminoácidos sulfurados, determinadas em porcos de engorda que tinham sido injetados por via intramuscular com uma solução salina ou uma solução de lipopolissacarídeo (LPS) de E. coli (Kim et al., 2012)
Manipulação do percurso dos eicosanóides
Um dos efeitos mais importantes da estimulação do sistema imunitário é o aumento da produção de eicosanóides como a prostaglandina E2 (PGE2) e o leucotrieno B4 (LTB4), que induzem a anorexia associada a infeções (Wymann e Schneiter, 2008) e controlam a duração da inflamação, respetivamente (Devchand et al., 1996). A produção de mediadores eicosanóides na célula e membranas nucleares é iniciada através da ação da fosfolipase A2 ou C que convertem, respetivamente, os fosfolípidos e os diacetilglicerois da membrana, em ácidos araquidónicos. Os ácidos araquidónicos são por sua vez convertidos em PGE2 ou LTB4 pelas ciclooxigenase e lipooxigenase, respetivamente (Folco e Murphy, 2006; Wymann e Schneiter, 2008; Kalinski 2012). Os ácidos gordos Omega-3 são conhecidos por reduzir de forma competitiva a produção de eicosanóides pró-inflamatórios, agindo como como substratos para as enzimas ciclooxigenase e lipooxigenase, enquanto que os antioxidantes, como a vitamina E e o selénio, reduzem os danos nos tecidos, atenuando a fagocitose associada as espécies reactivas ao oxigéno. A suplementação com boro reduz a resposta inflamatória ao unir as proteases produzidas durante a fagocitose (o mecanismo detalhado pode ser encontrado em Kim et al., 2013).
Conclusão
Os porcos alojados numa exploração comercial estão expostos, de forma consistente, a infeções (sub)clínicas e fatores de stress, e as respostas sistémicas para estes desafios reduz significativamente o potencial de crescimento dos porcos. A resposta imunitária afeta o rendimento do porco de duas formas diferentes: altera a partição de nutrientes e produz respostas neurológicas à infecção, como a anorexia. Como possiveis soluções é sugerida a suplementação com os aminoácidos que mais são utilizados durante a resposta sistémica para contrariar a alteração da partição de nutrientes, e a suplementação de nutrientes que reduzem a produção de mediadores eicosanóides que, por sua vez, provocam uma resposta neurológica à infecção.
*Alguns aspetos deste artigo foram publicados originalmente em “Impact of the systemic response to stressors and subclinical and clinical infection on intestinal barrier function and growth in pigs” em “Manipulating Pig Production XIV (eds. J.R. Pluske and J.M. Pluske), pp. 62-76, Australasian Pig Science Association.

