

A diarreia pós-desmame é uma das principais causas de perdas económicas na produção suína. Caracteriza-se por uma elevada incidência de diarreia e uma redução do crescimento atribuíveis a estirpes enterotoxigénicas de E. coli. Eventos como um desmame precoce e brusco, entre os 17 e os 28 dias de idade, geram um enorme stress, o que comporta uma perturbação na microbiota intestinal, na fisiologia e na função imunitária da mucosa. A resposta imunitária da mucosa intestinal é uma das áreas que tem tido mais atenção nos últimos 5 anos. O epitélio da mucosa é formado pelas células epiteliais, também conhecidas como enterócitos, que cobrem o tracto gastrointestinal. A sua saúde não é apenas importante para o crescimento e para o desenvolvimento do leitão devido à sua função de secreção e absorção mas também porque formam o maior órgão imunitário do porco e são a primeira resposta ante os microorganismos no intestino.
As células caliciformes que fazem parte da mucosa segregam muco e mucinas que fornecem a barreira inicial (figuras 1 e 2) (Varum et al. 2010 e Pelaseyed et al. 2014). As células epiteliais da mucosa também fornecem mucinas a esta barreira (Maynard et al. 2012 y Chase et al. 2012). Esta barreira também contém péptidos antimicrobianos (AMP) produzidos pelos enterócitos (figura 2). Estos AMP são induzidos pela microbiota do tracto intestinal e pelas células natural killer (NK) e LTi (figura 2) (Chase et al. 2012). Os AMP (também conhecidos como péptidos de defesa do hospedeiro, HDPs) são uma importante familia de moléculas que ajudan a formar uma barreira química para limitar a infecção nas superficies epiteliais e atacar as bactérias invasoras. Em porcos foram identificados e caracterizados mais de 30 HDPs (Maynard et al. 2012). Os AMPs são péptidos catiónicos relativamente pequenos e que se encontram, basicamente, nas superfícies mucosas e nas células fagocitárias. A sua estrutura e actividade antimicrobiana é variável e podem diferenciar-se funcionalmente. A concentração de alguns HDPs aumenta como resposta à inflamação ou infecção microbiana. A IgA secretora (sIgA) produz-se quando se segrega IgA dimérica por parte das células plasmáticas da lâmina própria e se transporta à superfície mucosa das células epiteliais (figura 2) (Maynard et al. 2012). A IgA secretora tem um papel importante na imunidade da superfície da mucosa ao aglutinar os agentes infecciosos, dificultando a sua aderência às células epiteliais e neutralizando toxinas. A camada mucosa interna juntamente com os AMP e as sIgA formam uma "zona letal" que poucos agentes patogénicos ou comensais podem penetrar (figura 2). Esta "zona letal" juntamente com as uniões estreitas, que são tecidas pelos enterócitos, forma uma "barreira" contra a E. coli e contra outros agentes patógénicos entéricos.

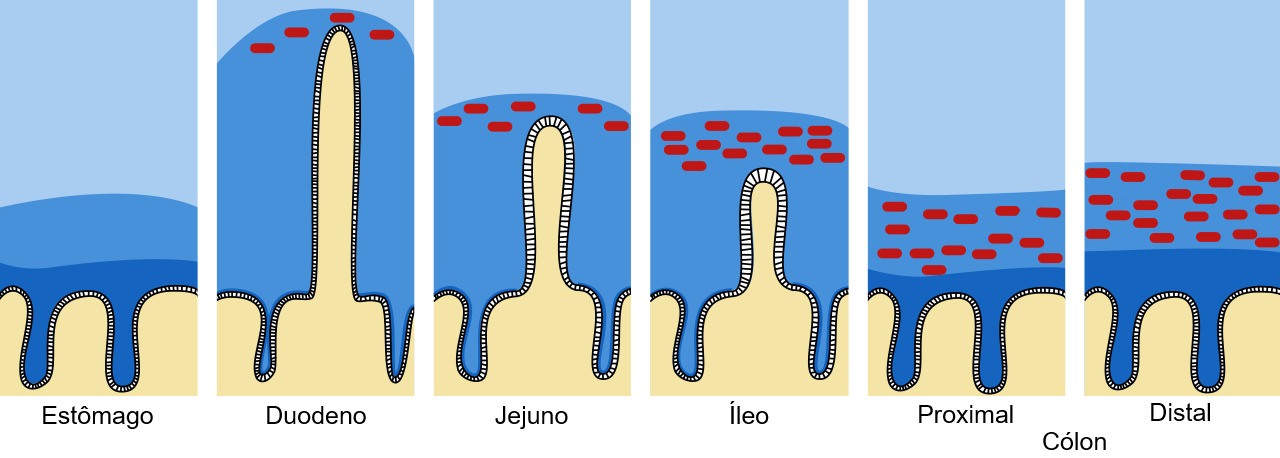
Figura 1. Barreira de muco e mucina no tracto intestinal. O muco é apresentado a azul escuro e as bactérias a encarnado. Adaptado de Pelaseyed T. et al. (2014). Immunological Reviews, 260(1), 8–20.
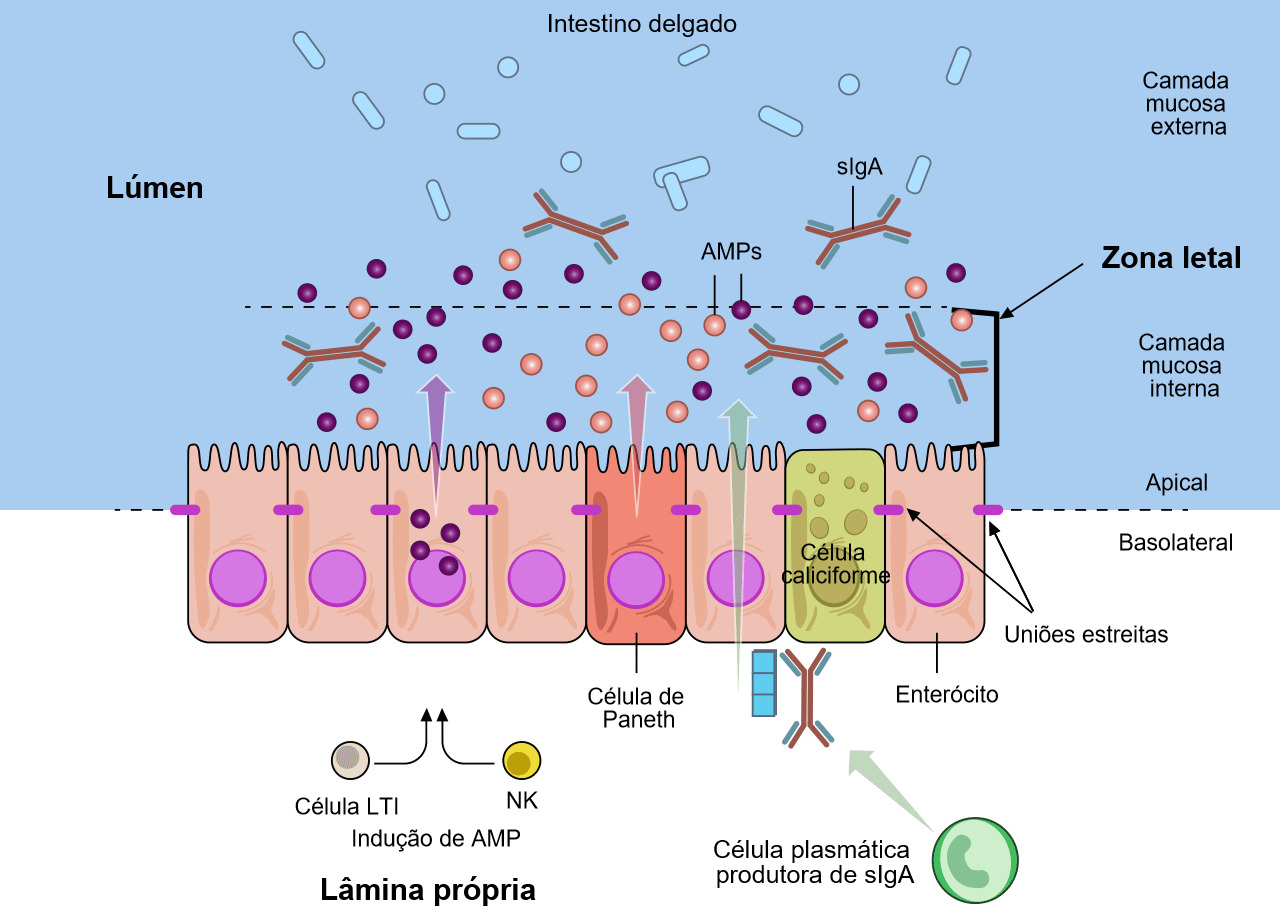
Figura 2. As defesas da mucosa do tracto gastrointestinal. Várias sub-populações de células epiteliais do intestino (IECs) estão integradas numa camada unicelular contínua que se divide numa região apical e nuna basolateral pelas uniões estreitas. Os enterócitos e as células especializadas de Paneth detectam a microbiota para induzir a produção de péptidos antimicrobianos (AMPs). As células caliciformes produzem mucina, que se organiza num gel denso de proteoglicanos muito entrelaçado, que forma uma camada interna aderente e uma camada externa menos entrelaçada. A camada externa está muito colonizada pelos constituintes da microbiota. A camada mucosa interna está muito impermeabilizada contra a colonização ou penetração bacteriana devido à sua grande concentração em AMPs bactericidas, bem como devido às IgA secretoras específicas (sIgA), que se deslocam da superfície basolateral, onde estão ligadas por um receptor à camada mucosa interna. Como resposta à microbiota, as células linfóides inatas, as células indutoras de tecido linfóide (LTi) e as células natural killer (NK), produzem citoquinas que estimulam a produção de AMP e mantêm a barreira epitelial. Adaptado de Maynard et al., Nature 489, 231–241 2012
Há outros componentes da resposta imunitária que também podem ser importantes na protecção contra vários tipos de infecção na superfície mucosa. No porco, os neutrófilos podem migrar massivamente para o lúmen intestinal como resposta a complexos antigene-anticorpo e matar as E. coli. A chegada de neutrófilos ao lúmen do intestino e a sua posterior destruição implica um aumento da concentração de lactoferrina, lisozima e proteínas catiónicas. Estas substâncias também podem contribuír para a imunidade contra infecções bacterianas do intestino (Maynard et al. 2012).
O desmame é um evento stressante para o leitão. Um desmame antes das 3 semanas implica efeitos negativos a longo prazo sobre o sistema imunitário e sobre o desenvolvimento da mucosa. O desmame precoce aos 17-21 dias gera um aumento prolongado das hormonas do stress, do factor libertador de corticotropina (CRF) e dos mastócitos, o que produz uma disfunção da mucosa intestinal (Chase et al. 2012).
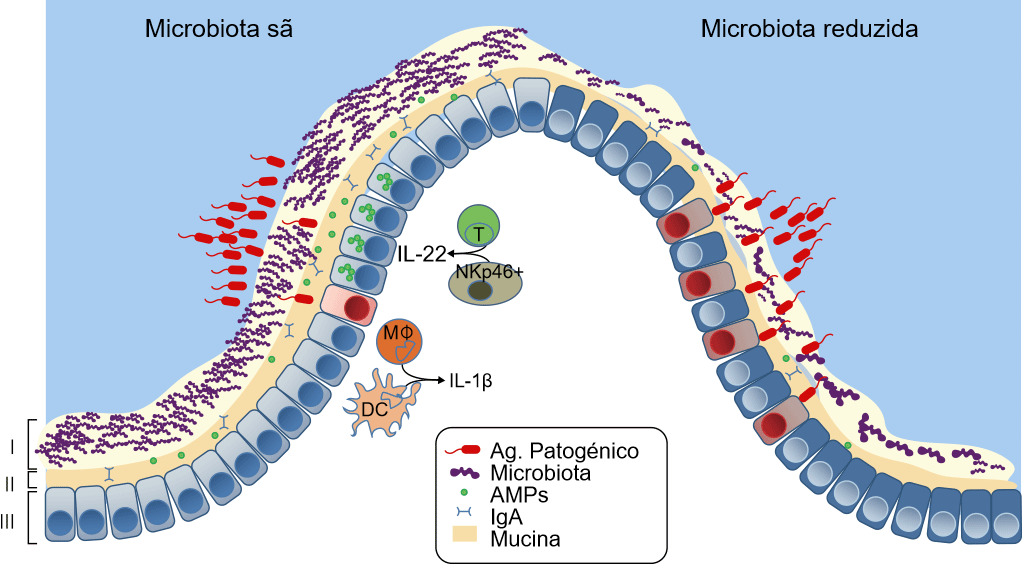
Figura 3. Defesas de uma mucosa sã ou com disbiose. A microbiota intestinal fornece três níveis de protecção contra a infecção entérica. (I) A microbiota satura as zonas de colonização e compete pelos nutrientes limitando a associação dos agentes patogénicos com o tecido do hospedeiro. (II) Zona letal - Os micróbios comensais preparam a barreira imunitária ao induzir a expressão da mucina, imunoglobulina A (IgA) e péptidos antimicrobianos (AMPs) que também previnem o contacto dos agentes patogénicos com a mucosa do hospedeiro. (III) Finalmente, a microbiota melhora a resposta imunitária contra os agentes patogénicos invasores. Isto consegue-se potenciando a expressão de IL-22 pelas células T e pelas NK, o que aumenta a resistência epitelial contra a infecção, bem como iniciando a secreção de IL-1b pelos monócitos intestinais (MK) e pelas células dendríticas (DCs) e isto, por sua vez, activa o recrutamento de células inflamatórias para a zona de infecção. Se a microbiota está ausente há menos concorrência, menos resistência da barreira e menos defesa imunitária contra a invasão de agentes patogénicos. Khosravi e Mazmanian, Current Opinion in Microbiology 2013, 16:221–22
O stress de ser separado da sua mãe, a idade ao desmame, a mistura num novo ambiente e a retirada repentina do leite materno alteram o ecossistema microbiano intestinal, reduzem as defesas ante a entrada de agentes patogénicos e aumentam o risco de doença, em particular, da diarreia pós-desmame (Fouhse et al. 2016). A diarreia pós-desmame caracteriza-se pela redução das bactérias comensais, como Lactobacillus spp, e pelo aumento das E. coli patogénicas (Fouhse et al. 2016). Isto gera uma disbiose — a perda das bactérias boas com um sobrecrescimento dos organismos nocivos como a E. coli (Fouhse et al. 2016 e Khosravi et al. 2013) (figura 3). Contudo, a disbiose não é apenas a perda da microbiota, também implica a redução da "zona letal" — a camada de muco fica menos espessa e a quantidade de sIgA e AMP reduz-se notavelmente, o que faz com que a barreira se debilite, permitindo que as E. coli enterotoxigénicas entrem em contacto com a mucosa e causem doença. Um dos factores principais que produzem disbiose e diarreia pós-desmame é a redução da ingesta de água e ração (Fouhse et al. 2016). A anorexia pós-desmame e a desidratação geram alterações estruturais no intestino, incluindo atrofia das vilosidades e hiperplasia das criptas, o que também leva à disfunção da barreira intestinal (McLamb et al. 2013). Os desmames precoces com menos de 20 dias de idade comportam uma deterioração da resposta imunitária inata com um número reduzido de neutrófilos e citoquinas inflamatórias que são necessários para controlar as infecções de E. coli pós-desmame (McLamb et al. 2013). Para piorar as coisas, a redução de bactérias produtoras de ácido láctico (Lactobacillus) durante o desmame faz com que aumente o pH intestinal, aumentando a susceptibilidade a doenças já que o pH baixo é bactericida para E. coli (Fouhse et al. 2016). As investigações destinadas a aumentar os micróbios simbióticos e reduzir os agentes patogénicos oportunistas centram-se na redução da disbiose.