

A Escherichia coli (ETEC) enterotoxigénica continua a ser uma das causas principais de doença e morte em porcos recém-nascidos e recém-desmamados. Os porcos que desenvolvem diarreia após o desmame (diarreia pós-desmame; DPD) costumam fazê-lo entre os dias 3 a 10. As estirpes de ETEC que causam DPD possuem 2 tipos de factores de virulência, adesinas e enterotoxinas e ambos são essenciais para a expressão clínica da DPD. Os isolados de ETEC suína produzem qualquer das cinco adesinas diferentes, todas elas são fímbrias (ou pili): K88 (F4), K99 (F5), 987P (F6), F41 (F7) e F18. As adesinas fimbriais K88 e F18 podem ter diferentes formas antigénicas, a mais frequente, K88, inclui as variantes K88ab, K88ac e K88ad. As adesinas fimbriais fixam-se a receptores específicos na extremidade dos enterócitos do intestino delgado, o que permite às bactérias colonizar a superfície celular e excretar toxinas. As toxinas principais produzidas pelas estirpes de ETEC que causam DPD são as termolábeis (LT), as termoestáveis tipo A (STa), as termoestáveis tipo B (STb) e (ou) a enterotoxina termoestável 1 enteroagregativa de E. coli (EAST1). A enterotoxina Stx2e (toxina shiga), também conhecida como factor da doença dos edemas, causa as lesões associadas à doença dos edemas em porcos (ver resumo na tabela 1).
Deve ser destacado que a subunidade B de LT (LTB) tem propriedades imunomoduladoras que podem produzir apoptose (morte celular) das células T CD8 positivas, activação das células B e alteração da secreção de citoquina pelos monócitos. As propriedades adjuvantes da LTB foram evidenciadas num estudo em porcos desmamados que mostrou uma maior protecção frente a ETEC DPD mediante uma vacina viva de E. coli com genes para LTB e K88ac em comparação com uma estirpe isogénica sem o gene para LTB. A estirpe que apenas possuía LTB não conseguiu gerar uma imunidade protectora. Por outro lado, as toxinas STa e STb são moléculas pequenas e consideradas pouco imunogénicas e estes antigénios por si só não podem induzir a resposta imunitária anti-STa e anti-STb requeridas para uma protecção efectiva frente a DPD no porco.

Tabela 1. Resumo dos factores de virulência de ETEC associados a DPD (reproduzido de Zhang, 2014).
| I. Adesinas | Referências | |
| Adesinas fimbriais | Morfología (diâmetro) e sub-unidade estrutural (kDa) | |
| K88 (F4) | Fibrilar (2,1 nm), FaeGa(27,6)/FaeCb(16,9) | Bakker et al., 1991 |
| K99 (F5) | Fibrilar (4,8 nm), FanCa(16,5)/FanFb(31,5) | Isaacson, 1977; Bakker et al., 1991 |
| 987P (F6) | Fimbrial (7,0 nm), FasAa(23,0)/FasFb(17,5)/FasGb(40) | Isaacson & Ritcher, 1981; Edwards et al., 1996 |
| F41 (F7) | Fibrilar (3,2 nm), Fim41aa(29,5) | deGraaf, 1982 |
| F18 (F107; 2134P; CFA8813) | Fibrilar (3-4 nm), FedAa(15,1)/FedFb(30,3) | Imberechts et al., 1996; Nagy et al., 1997 #17795; Smeds et al., 2001 |
| Adesinas não fimbriais | ||
| AIDA-I | Proteína autotransportadora, AidA (79,5c) | Benz & Schmidt, 1992; Benz & Schmidt, 2001 |
| ppa | Proteína da membrana externa, Paa (27,6) | Batisson et al., 2003 |
| EAE | Proteína da membrana externa, EaeA (94-97) | Jerse & Krapper, 1991 |
| II. Toxinas | Gene, proteína de toxina madura | |
| LT | eltAB, 1:5 AB holotoxina (86 kDa) | Sixma et al., 1991; Streatfield, 1992 |
| STa | estA, péptido 18 aminoácidos (2kDa) | Dreyfus et al., 1983 |
| STb | estB, péptido 48 aminoácidos (5,1 kDa) | Lee et al., 1983; Dreyfus et al., 1992 |
| EAST1 | astA, péptido 38 aminoácidos (4,1 kDa) | Nataro et al., 1987 |
| Stx2e | stx2e, 1:5 AB toxin (70 kDa) | Pierard et al., 1991 |
a: subunidade estrutural maior
b: subunidade estrutural menor e/ou da ponta fimbrial
c: adesina madura AIDA-I detecta-se como 100 kDa mediante SDS-PAGE
Nem todas as estirpes ETEC possuem a fímbria F4 e isto explica, em parte, algumas das variações vistas entre explorações em DPD. No entanto, a vacinação de uma porca durante a gestação leva à secreção de anticorpos antigénio-específicos no colostro e no leite, que podem proteger os leitões antes do desmame contra a infecção por ETEC. No entanto, o desmame causa a perda dos anticorpos e IgG do leite, o que parece contribuir para a maior susceptibilidade dos porcos às infecções entéricas por E. coli após o desmame e, portanto, é necessária uma resposta imunitária activa das mucosas para proporcionar protecção. Em concreto, para induzir uma resposta imunitária das mucosas que seja protectora é requerida a activação do sistema imunitário da mucosa do intestino delgado, especificamente das placas de Peyer do jejuno, para provocar uma resposta IgA ou IgM específica para antigénios F4. Tal como se observa na figura 1, no entanto, é requerido certo tempo após o desmame para que os níveis de IgA aumentem em resposta a qualquer desafio patogénico.
Devido a que as estirpes de ETEC são, de longe, a causa mais comum e importante de DPD, estas E. coli patogénicas têm sido o objectivo principal do desenvolvimento de vacinas. A vacina frente a DPD esteve em desenvolvimento durante muitos anos, no entanto, tal como explica Zhang (2014), é complicado produzir uma vacina rentável contra DPD. As enterotoxinas determinam a virulência da DPD, pelo que devem ser incluídas como componentes vacinais inclusive quando LT, STa e STb são potencialmente tóxicas e não podem ser utilizadas como antigénios seguros. Portanto, uma vacina eficiente frente a DPD deveria induzir imunidade anti-adesina frente às fímbrias K88 e (ou) F18 e também imunidade antitoxina pelo menos contra LT, STa e STb. Estas fímbrias e toxinas são genética e imunologicamente heterogéneas e, portanto, a imunidade induzida por uma fímbria ou toxina apenas proporciona protecção contra as estirpes ETEC que expressem a mesma fímbria ou toxina, mas não pode haver protecção cruzada contra estirpes ETEC que expressem fímbrias ou toxinas diferentes.
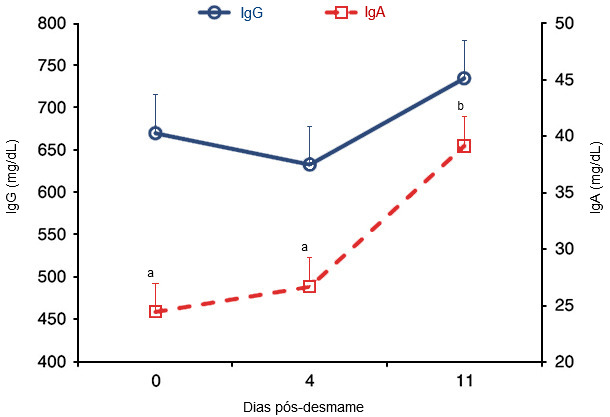
Figura 1. Concentrações de IgG e IgA (mg/dL) plasmáticas nos dias 0, 4 e 11 após o desmame. A concentração de IgA foi mais elevada (P < 0,001) no dia 11 que no dia 0 ou 4 após o desmame. [Os porcos foram desmamados no dia 0 e expostos a E. coli F18 nos dias 1, 2 e 3 após o desmame. O plasma foi recolhido no dia 0 antes dos leitões terem sido transferidos para as baterias] (Sugaharto et al., 2014).
Além disso, os porcos de diferentes origens genéticas expressam receptores diferentes, que reconhecem e aos quais se derem fímbrias específicas expressas por diferentes estirpes de ETEC. Se os porcos forem imunizados oralmente com uma vacina fimbrial, apenas os porcos que expressem receptores específicos para esta fímbria desenvolverão uma resposta imunitária efectiva. Ainda assim, uma vacina eficiente frente a DPD deveria induzir uma imunidade protectora anti-adesina frente às fímbrias K88 e (ou) F18 mas também imunidade antitoxina frente a LT e ST. O uso de um antigénio toxóide de fusão multiepitope, MEFA, (que inclui anticorpos neutralizantes frente a toxinas) permitiu a inclusão potencial de múltiplos antigénios de adesão e de toxinas num único produto para o desenvolvimento de vacinas seguras e de amplo espectro real frente a DPD, no entanto, são necessárias mais experiências antes da sua comercialização.
Na prática, e para conseguir ter anticorpos adquiridos activamente preparados no momento do desmame, os leitões deveriam ser imunizados entre os 10 dias e as 2 semanas antes do desmame. No entanto, o nível necessário de anticorpos maternais passivos nos leitões lactantes deve ser suficientemente grande para proteger contra a diarreia neonatal mas suficientemente bajo para que não comprometa a indução de anticorpos específicos frente a DPD. A vacinação individual dos porcos ou uma imunização oral no momento do desmame (p.e., adicionando produtos vacinais na ração ou na água de bebida) são opções pelas quais se pode gerar imunidade.


