

A resposta humoral específica começa quando um linfócito B naïve reconhece o seu antigene específico nos tecidos e órgãos linfóides secundários (ver "Reconhecimento de agentes patogénicos").
Cada linfócito B tem um receptor de antigene (BCR: Receptor de células B), uma imunoglobulina de superfície (IgM ou IgD), que se une a domínios específicos do antigene chamados determinantes antigénicos ou epítopes.

A especificidade do sítio de união antigénica depende dos genes que codificam os domínios variáveis (VH e VL); nos mamíferos a recombinação e/ou conversão génica podem gerar um grande número de genes e, em consequência, muitos BCRs distintos, capacitando o animal para reconhecer quase todos os antigenes externos.
Após o reconhecimento, cada linfócito B específico activa-se, realiza uma proliferação clonal e diferencia-se numa célula plasmática (ou Célula Secretora de Anticorpos, ASC) capaz de sintetizar e secretar imunoglobulinas (anticorpos), específicas para o seu epítope.
As células T ajudam a optimizar a resposta humoral específica.
Inicialmente, as células B secretam IgM sob a forma monomérica e com uma especificidade antigénica idêntica.
Em relação ao tipo de secreção de anticorpos, os antigenes classificam-se como dependentes ou independentes das células T, consoante necessitem, ou não, da ajuda das células T para iniciar a resposta dos anticorpos.
Para certos antigenes, os chamados independentes das células T (por exemplo lipopolissacáridos bacterianos, flagelina, polissacáridos de pneumococos, etc.) só podem induzir anticorpos IgM com uma afinidade modesta e sem células B de memória.
Quase todos os antigenes externos são dependentes das células T e necessitam, para a activação completa dos linfócitos B, um sinal adicional (citoquina) procedente dos linfócitos T colaboradores específicos que reconheçam o mesmo antigene (cooperação das células T e B).
Neste último caso, a célula B actua como uma célula apresentadora de antigenes, apresentando o antigene processado às células T colaboradoras que, quando é reconhecido, permite a mudança isotípica da Ig (comutação de classe de anticorpo) da IgM para imunoglobulinas de uma classe distinta, IgA, IgE ou IgG, dependendo da citoquina secretada pela célula T colaboradora, e envolvidas na estimulação da célula B (IL-4 ou IFNγ respectivamente). Outro sinal básico para a comutação do anticorpo é a interacção do CD154 (CD40L) no receptor CD40 das células B.
Sob esta "ajuda", as células B proliferativas têm uma hipermutação somática da imunoglobulina de superfície, pelo que passarão a estar dotadas de um isotipo específico. Estes linfócitos B são posteriormente seleccionados com base numa afinidade óptima da Ig com o antigene ao entrar em estreito contacto com as células dendríticas foliculares que apresentam o antígene. Só os linfocitos B com uma elevada afinidade do anticorpo para o antigene e que são capazes de o processar e apresentar, serão seleccionadas positivamente. Portanto, só estes últimos linfócitos B sobreviverão, proliferarão e diferenciar-se-ão em células plasmáticas, sintetizando e secretando anticorpos de uma única classe isotípica, com uma especificidade única e afinidade elevada, melhorando a capacidade de aderir ao antigene e, deste modo, neutralizar e destruir os agentes patogénicos.
Quando termina a resposta imunitária primária, algumas destas células B e células plasmáticas activadas e seleccionadas neste primeiro contacto, sobreviverão como células de memória de vida longa, prontas para serem activadas em futuras exposições ao mesmo antigene (resposta imunitária secundária) produzindo a mesma classe de anticorpos com um maior título e afinidade.
Em conclusão, na resposta humoral primária, inicialmente é secretada uma única classe de IgM sob a forma monomérica e com idêntica especificidade antigénica. Uns dias depois, devido à comutação isotípica, são secretadas classes distintas de Ig, por exemplo, IgG. A resposta humoral secundária induzida por uma exposição posterior ao mesmo antigene será caracterizada, principalmente, pela produção do isotipo de IgG mas com um título e afinidade mais elevadas (figura 1).
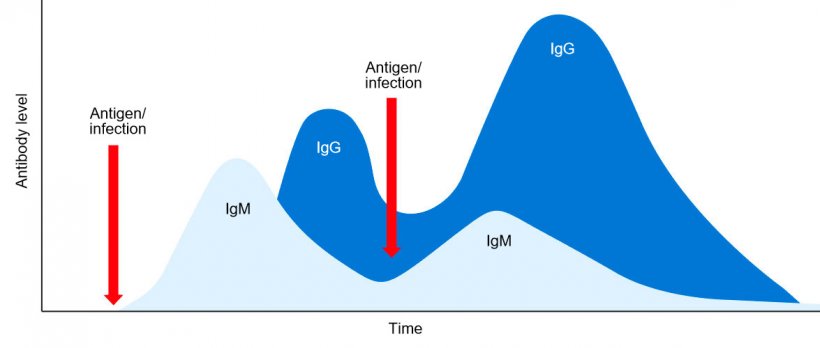
Figura 1: Cambio isotípico y respuesta inmune primaria y secundaria a exposiciones posteriores al mismo antígeno.
Os anticorpos podem actuar contra agentes patogénicos com diferentes mecanismos defensivos:
- Inibição da infectividade de agentes patogénicos unindo-se directamente a eles (neutralização, bloqueio por adesão, exclusão imunitária).
- Activação da via alternativa (opsonização) ou clássica do complemento com lise do agente patogénico.
- Ao cobrir o agente patogénico (ou células infectadas), os anticorpos permitem que as células acessórias, que reconhecem partes do fragmento cristalizável (Fc), ingiram e matem o agente patogénico por opsonização e fagocitose ou matem as células infectadas mediante ADCC ("Citotoxicidade Celular dependente de Anticorpos").
- Regulação da resposta imunitária através da interacção com os receptores Fc (FcRs).
Os FcRs expressam-se em muitas células efectoras da imunidade inata, como os mastócitos, os neutrófilos, os monócitos, os macrófagos e as células dendríticas. Concretamente, quando os FcγRs (receptores Fc para IgG) se unem a complexos imunitários, desencadeiam a activação das células inatas e a sua resposta efectora (fagocitose, explosão oxidativa, citotoxicidade, libertação de péptidos vasoactivos e quimioatraentes). Os FcRs das células dendríticas promovem a captura do antigene, o seu processamento e a sua apresentação às células T citotóxicas e colaboradoras.
Pelo contrário, a FcR inibitória para a IgG, expressa nas células B, está implicada na regulação da activação e desenvolvimento de células B e na sobrevivência de células plasmáticas.


